To tylko jedna z 20 stron tej notatki. Zaloguj się aby zobaczyć ten dokument.
Zobacz
całą notatkę

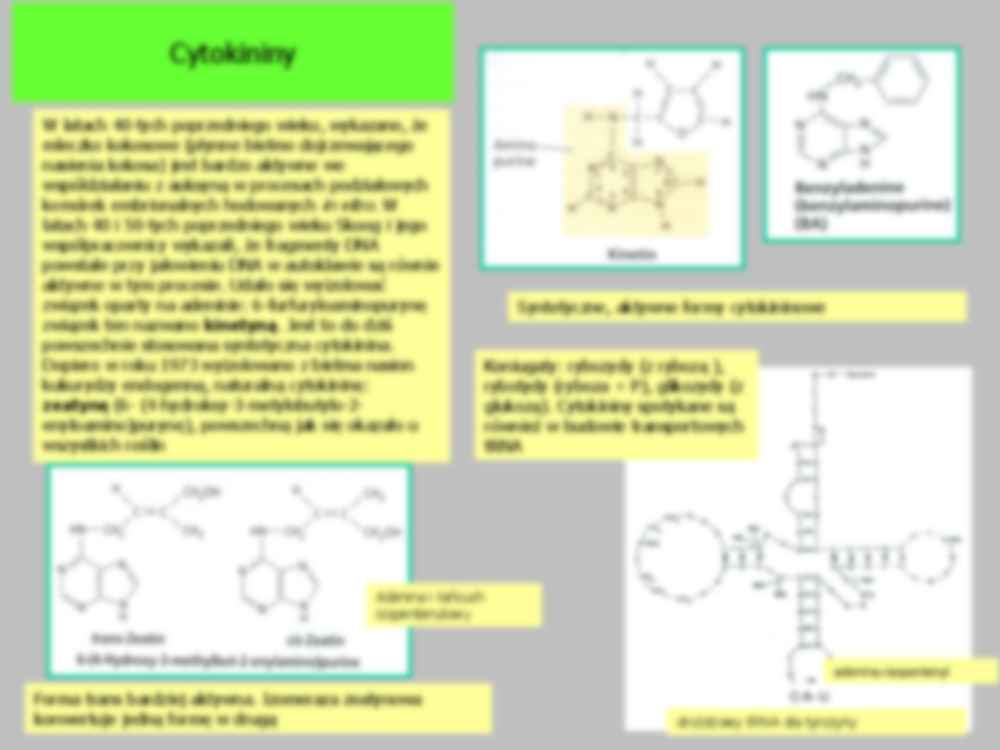
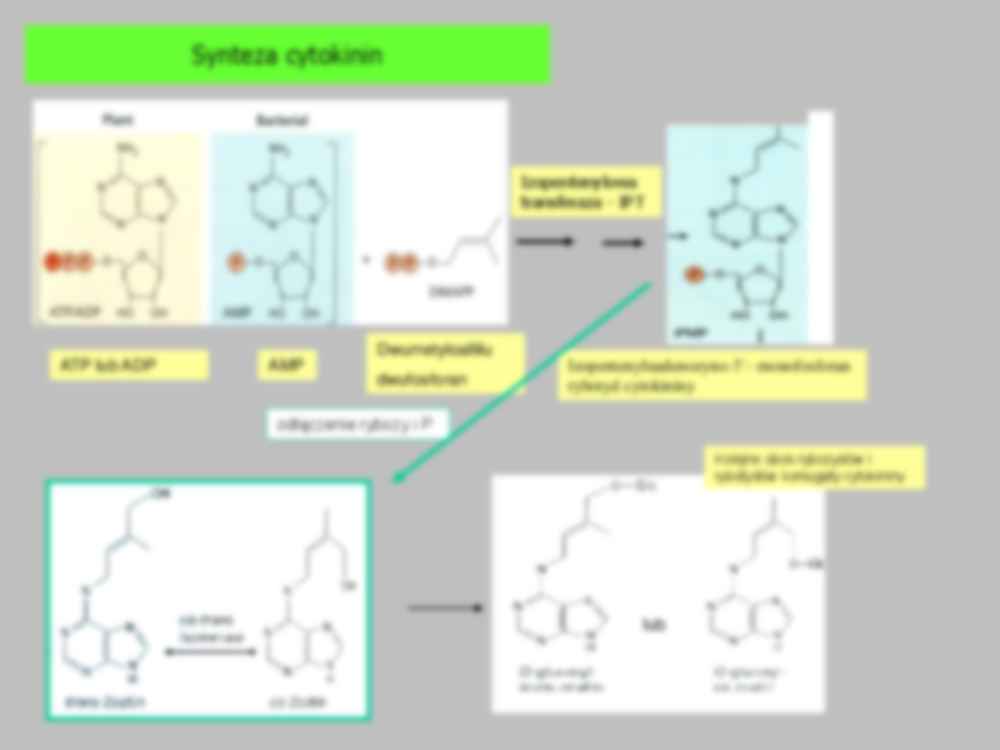
Cytokininy Cytokininy W latach 40-tych poprzedniego wieku, wykazano, że mleczko kokosowe (płynne bielmo dojrzewającego nasienia kokosa) jest bardzo aktywne we współdziałaniu z auksyną w procesach podziałowych komórek embrionalnych hodowanych in vitro. W latach 40 i 50-tych poprzedniego wieku Skoog i jego współpracownicy wykazali, że fragmenty DNA powstałe przy jałowieniu DNA w autoklawie są równie aktywne w tym procesie. Udało się wyizolować związek oparty na adeninie: 6-furfuryloaminopurynę związek ten nazwano kinetyną . Jest to do dziś powszechnie stosowana syntetyczna cytokinina. Dopiero w roku 1973 wyizolowano z bielma nasion kukurydzy endogenną, naturalną cytokininę: zeatynę (6- (4-hydroksy-3-metylobutylo-2- enyloamino)purynę), powszechną jak się okazało u wszystkich roślin Forma trans bardziej aktywna. Izomeraza zeatynowa konwertuje jedną formę w drugą Syntetyczne, aktywne formy cytokininowe drożdżowy tRNA dla tyrozyny adenina+isopentenyl C-A--U Koniugaty: rybozydy (z rybozą ), rybotydy (ryboza + P), glikozydy (z glukozą). Cytokininy spotykane są również w budowie transportowych tRNA Adenina i łańcuch izopentenylowy Synteza cytokinin Dwumetyloallilu dwufosforan ATP lub ADP AMP Izopentenyluadenozyno-5’- monofosforan rybotyd cytokininy Izopentenylowa transferaza - IPT odłączenie rybozy i P lub Kolejne obok rybozydów i rybotydów koniugaty cytokininy Rozkład cytokininy adenina 3-metylo-2-butenal Różna nadekspresja genu oksydazy cytokininowej TYTOŃ Przekrój przez merystem odmiany dzikiej tytoniu (A) i odmiany transgenicznej z nadekspresją genu oksydazy cytokininowej (B) Redukcja wielkości merystemu rośliny transgenicznej z nadekspresją genu oksydazy CK niski poziom aktywnej cytokininy A B W merystemie cytokinina niezbędna do stymulacji podziałów komórkowych 1. Czynnik transkrypcyjny STM indukuje syntezę izopentenylowej transferazy (IPT) umożliwiającej syntezę cytokininy 2. Czynnik transkrypcyjny WUS hamuje syntezę represora działania cytokininy (ARR5/ARR7) 3. Cytokininy wspólnie z regulatorem STM hamują działanie GA w kierunku różnicowania komórek Miejsce syntezy i transport Podstawowym miejscem syntezy cytokinin jest merystem apikalny korzenia i młode zarodki. Ostatnie badania wykazały że CK są syntetyzowane również przez młode, rozwijające się, liście i młode owoce Z korzeni CK transportowane są ksylemem, pod postacią
(…)
… – fosforylacja histydyny
1.
2.
3
Np.: indukcja ekspresji genu
odpowiedzialnego za cyklinę
CYC D3
Histydynowe białko transferujące P (AHP –
Arabidopsis histidine proteins) przeniesienie P z asparaginianu na
histydynę. AHP zwykle przemieszcza się
do jądra komórki
3.
2
Domena odbierająca z asparaginianem .
Odbiór P od histydyny
Regulator odpowiadający, ( reagujący )
(ARR – Arabidopsis response regulator)przeniesienie P z histydyny AHP na
asparaginian. Białka ARRs mogą działać
bezpośrednio jako czynniki transkrypcyjne
H
Asp
1.
Arabidopsis - cytokinina łączy się ze
swoim receptorem błonowym –
CRE(zwykle występuje jako dimer) (do
domeny przyjmującej zwanej CHASE.
Podobnie funkcjonują obecne u
Arabidopsis AHK2 i AHK3 (też
receptory CK)
2.
Połączenie CK indukuje fosforylację
histydyny (domena przenosząca) i dalej…
... zobacz całą notatkę
Komentarze użytkowników (0)