To tylko jedna z 23 stron tej notatki. Zaloguj się aby zobaczyć ten dokument.
Zobacz
całą notatkę


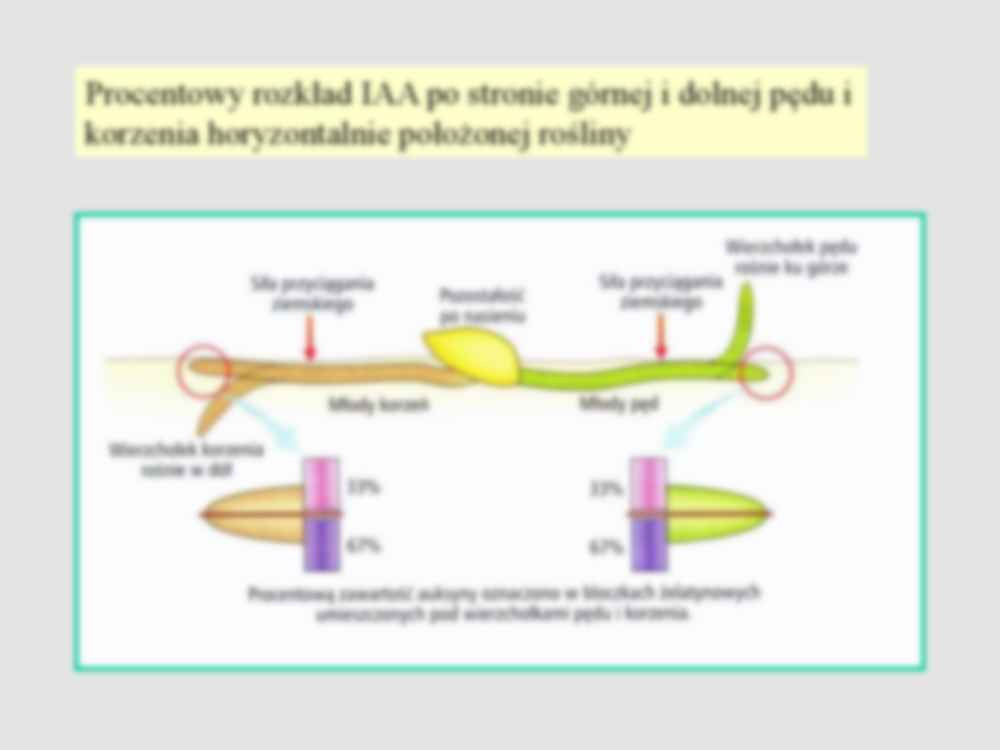
TROPIZMY – ruchy wzrostowe, wywołane przez bodźce ukierunkowane Wzrostowe Ukierunkowan e Fototropizm – ruch wzrostowy zależny od kierunku padania światła; fototropizm dodatni pędu (do światła), fototropizm ujemny korzenia (od światła) tropizmy korzeń pęd Więcej IAA po stronie dolnej – szybszy wzrost Zbyt duże stężenie IAA po stronie dolnej – szybszy wzrost strony górnej Ruchy związane z nierównomiernym wzrostem różnych stron organu, skierowane do (+) lub od (-) źródła działania bodźca fototropizm Geotropizm (grawitropizm) Hormonalna hipoteza Chołodnego-Wenta Transport IAA na stronę nie naświetloną Transport iAA na stronę dolną Procentowy rozkład IAA po stronie górnej i dolnej pędu i korzenia horyzontalnie położonej rośliny Fototropizm (historia) 1 2 3 4 1. Konieczność obecności wierzchołka 2- 3 Możliwość spływu po stronie nie naświetlonej 4. Wygięcie można zaindukować bez światła Uruchomienie genu GH 3 przez IAA. Tam gdzie dużo IAA tam więcej aktywnego genu. Reakcja barwna z wykorzystaniem genu reporterowego GUS. Gen GUS dołącza się do innego genu tutaj do genu GH 3. Gen GUS uruchamia się równocześnie z genem do którego był podłączony GUS – enzym – beta glukouronidaza,. Generująca barwny produkt z bezbarwnego substratu w roztworze w którym zanurzone są badane tkanki Kierunek padania światła Fototropina - NPH1 Receptor światła niebieskiego: flavoproteina NPH1 –fototropina (120 kDa) – kinaza samofosforulująca się. Nie wiadomo jaki jest mechanizm NPH1- P na lateralny transport IAA. Zakłada się pewną rolę w procesie kryptochromów CRY1 i CRY2 Nie ma zależności: silniejsze światło – lepsze wygięcie fototropowe Przy mocniejszym świetle obserwuje się negatywne wygięcie (owies) lub brak wygięcia np.: soczewica, czy rzepak. Brak wygięcia może być w dość szerokim zakresie natężenia światła np.: słonecznik czy żyto. Dopiero jeszcze mocniejsze światło daje drugie pozytywne wygięcie fototropowe Przy niskim świetle obserwuje się gradient NPH1- P na delikatniejszym wierzchołku. Przy pewnej sile światła ten gradient się wyrównuje (brak wygięcia). Mocniejsze światło indukuje powstanie gradientu NPH1 poniżej wierzchołka (drugie pozytywne wygięcie) Trudności w określeniu mechanizmu fototropowego u dużych roślin zielonych. Dodatkowe źródła IAA (młode liście, pączki boczne). Często
(…)
…, a także umożliwiającej dimeryzację ARFów
Zidentyfikowano różne ARF-y u Arabidopsis thaliana
bogaty w glutaminę, serynę, leucnę (6,7,8,19),
syrenę, leucynę prolinę (1,2,3,4,9,11,12,14,15,18,20,21,22)
serynę., leucnę lub glicynę (3,13)
seryne, prolinę, leucynę z inną sekwencją w DBD (10,16)
bogaty w serynę z inna sekwencją w DBD (17)
niekompletny (23)
Dwie drogi (dwa receptory) indukowania zakwaszania ściany
komórki…
… receptor TIR i
kompleks SCF to:
•
•
•
Aux/IAA – białka tych genów
stanowią represory genów
wczesnych
SAUR – możliwe, że produkty
tych genów są istotne w
tropizmach
GH3 - kodują białko o MW= 70
kDa o funkcji nieznanej
Do niedawna do wczesnych genów
indukowanych przez IAA
zaliczano jeszcze:
•
•
Geny kodujące s-transferazę
glutationową, odpowiedzialną
za transport do wakuoli różnych
trucizn i niektórych…
... zobacz całą notatkę
Komentarze użytkowników (0)